TEXT
(Click on the picture below to open the link)

RELATED TOPICS
ANIMAL NOMENCLATURE
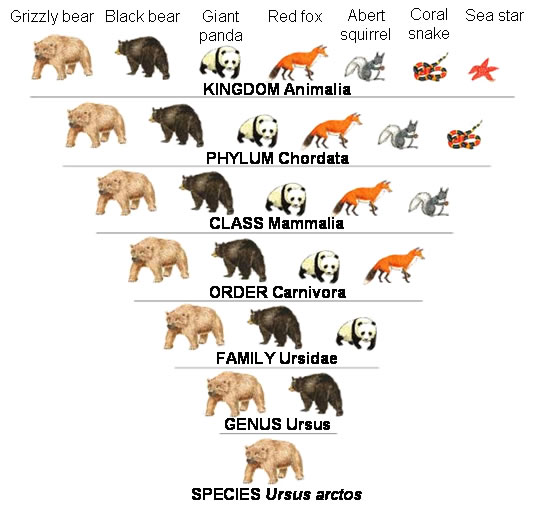

TYPES OF “SAUR”
Dinosaur: From the Greek words “Deinos” (terrible) and “Sauros” (lizard).
OTHER “SAURS”: Types of -saurs that are not dinosaurs
GEOLOGICAL TIME PERIODS
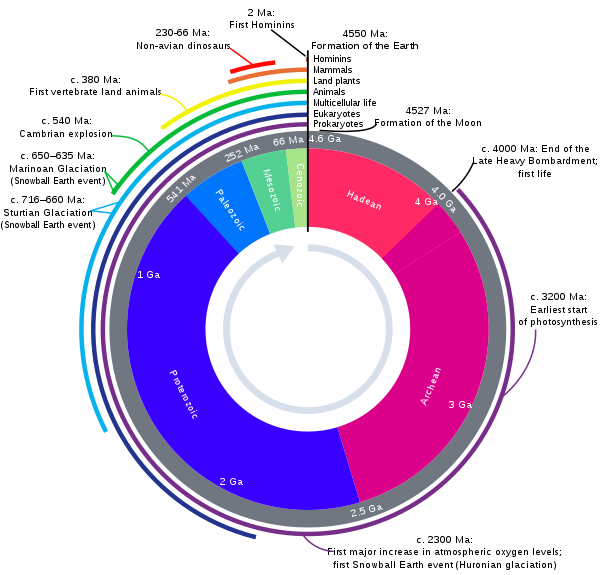
LEXICON
(THIS TEXT IS REPRODUCED HERE FOR THE ONLY PURPOSE OF HELPING THE READER WHO IS STUDYING ENGLISH FIND RAPIDLY THE DEFINITIONS OF UNFAMILIAR WORDS – FOR THE FULL ARTICLE, SEE THE WEBSITE)
In the early 19th century, long before the discovery of the dinosaurs, scientists and the public alike were faced with the realization that strange beasts, wholly extinct, were once populating Earth’s ancient oceans. In no small part, this realization was through the discovery of the first plesiosaurs (and ichthyosaurs) along the Dorset coast of England in the seaside town of Lyme Regis. There was this large marine reptile resembling a large sea turtle, but with four evenly shaped flippers and looking as though a large snake had been pulled through its carapace. It was soon to be named scientifically Plesiosaurus, in reference to its greater similarity to living reptiles than the Ichthyosaurus (Figure 1). While the Ichthyosaurus was relatively easily understood as a fish-shaped reptile descended from land-living ancestors, the Plesiosaurus was beyond comprehension, even though incomplete skeletons had been unearthed already in the early 18th century. Plesiosaurs seemed so alien that the first complete skeleton, discovered by the famed Mary Anning a little more than 200 years ago (Figure 1A), was considered a fake by the leading anatomist of the day, the Baron Georges Cuvier in Paris. Only study of the original specimen convinced him of the authenticity of this animal but reinforced his seminal insight that there is extinction.
Main text
What intrigued Cuvier most, and still offers a challenge to evolutionary developmental biologists today, was the incredibly long neck (Figure 1A), consisting of over 40 (or more) vertebrae, almost twice as many as any other tetrapod. The tail, by contrast, was short, unlike in most other reptiles. This combination of a short tail and an unreasonably long neck precipitated another famous episode in the history of paleontology some decades later. Reconstructing the most long-necked of plesiosaurs, an elasmosaur, American paleontologist and zoologist Edward Drinker Cope had mounted its skull on the tip of its tail (Figure 1C). Apparently, he too could not cope with the notion of an animal having over 70 neck vertebrae. When this error was repeatedly noted in public by his competitor Othniel Charles Marsh, the ‘bone wars’ ensued, which later played out in the race over dinosaur discoveries in the Wild West.
Figure thumbnail gr1
Figure 1190 years of plesiosaur life restorations.
Show full caption
View Large ImageFigure ViewerDownload Hi-res imageDownload (PPT)
Returning to the first plesiosaur skeleton, the flippers presented a puzzle as well. Their anatomy is not very different from the front limbs of whales and sea turtles, but the hind flippers, which are absent in whales and small in sea turtles, look very much like the front flippers (Figure 2). In fact, on occasion, front flippers have been mixed up with hind flippers in mounted skeletons in museums, in the tradition of Cope. The similarity in shape of fore- and hindlimbs is also a unique feature among tetrapods, suggesting that all limbs served exactly the same function.
Figure thumbnail gr2
Figure 2Plesiosaur fossils.
Show full caption
View Large ImageFigure ViewerDownload Hi-res imageDownload (PPT)
Popular interest in the sea monsters from Lyme Regis was such that, in 1830, it led to an artistic visualization of plesiosaurs as living animals, the beginnings of paleoart. The genre started with a bang, as a loose-leaf lithograph entitled Duria Antiquior (a more ancient Dorset; Figure 1B). This iconic depiction of a plesiosaur attacked by an ichthyosaur surrounded by a marine menagerie in an Early Jurassic sea is the archetype of many such illustrations, immortalized by Jules Verne in his Journey to the Center of the Earth from 1864 (Figure 1D). Paleoart, even in its earliest version, derives its scientific relevance by visualizing paleobiological hypotheses, albeit more or less well supported and often only implicitly. However, such visualizations sometimes inspire real-life scientific questions such as: How exactly did plesiosaurs use their four equally shaped flippers in locomotion, or why was the plesiosaur neck so incredibly long, and how flexible was it? These questions are at the center of much modern plesiosaur research and lend importance to the study of unique but extinct Bauplans such as that of plesiosaurs for our understanding of evolution.
A tale of three extinctions
Often called ‘the mother of all extinctions’, the end-Permian mass extinction 251.9 million years ago virtually emptied the oceans of life. A major pattern in the recovery is the colonization of the sea by different kinds of land-living, air-breathing amniotes. Prominent examples are the ichthyosaurs and the sauropterygians, of which the plesiosaurs are a subgroup. These lineages, collectively known as ‘Mesozoic marine reptiles’, are the geologically earliest examples of life transitioning from land back to the ocean. It is increasingly becoming clear that recovery from the catastrophe was extremely fast, and marine reptiles diversified within the first few million years of the Mesozoic. In fact, ever older finds dating to the earliest Triassic hint at a pre-Triassic origin of at least the ichthyosaurs but possibly also the sauropterygians.
Ichthyosaurs dispersed over the world’s ocean (Panthalassa), whereas sauropterygians were restricted to the precursor of the Mediterranean, the Tethys ocean; except for one lineage, the pistosaurs, of which plesiosaurs are a subgroup (Figure 3).
Figure thumbnail gr3
Figure 3Calibrated plesiosaur phylogeny.
Show full caption
View Large ImageFigure ViewerDownload Hi-res imageDownload (PPT)
In the Triassic, non-plesiosaur pistosaurs left scanty records not only in the Tethys but also in Siberia and in eastern Panthalassa. Like other sauropterygians, non-plesiosaur pistosaurs were propelled by their tail. However, they already show many similarities with plesiosaurs, including the skull architecture and the long neck. This is why non-plesiosaur pistosaurs have long been suspected to have given rise to the plesiosaurs in the Late Triassic, but good intermediate forms are missing to this day, due to the poor fossil record of marine reptiles in the last 25 million years of the Triassic. Only recently, evidence of the Triassic origin of plesiosaurs came to light in a clay pit in Germany in the form of Rhaeticosaurus. This skeleton of a small plesiosaur is the only one from the Triassic.
The second mass extinction, at the end of the Triassic, 201.4 million years ago, played an important role in plesiosaur evolution. It seemingly wiped out (or nearly so) most of the competition, including all other sauropterygians (Figure 3), thalattosaurs and giant ichthyosaurs. Then, in the earliest Jurassic, there is an abundance of small (<100 kg body mass) plesiosaurs from places like Lyme Regis (Figure 1A). This had suggested that plesiosaurs evolved very rapidly in the aftermath of the extinction event in the earliest Jurassic, just as their ancestors had done in the earliest Triassic. However, phylogenetic inference, corroborated by the relatively derived position of Rhaeticosaurus, reveals that plesiosaurs diversified before the end of the Triassic (Figure 3). In addition, isolated large plesiosaur bones from the Late Triassic represent animals that must have weighed several hundred kilograms and suggest that plesiosaur body size took a dip across the boundary.
Based on a largely European record from the first 20 million years of the Jurassic, plesiosaur diversification continued, and maximum body size gradually evolved to pre-extinction levels. Larger body size correlates with an increasingly macropredatory lifestyle (feeding on other marine reptiles) in some plesiosaurs: the iconic long neck became shorter, and the skull very large (Figures 2 and 3). This macropredatory body plan is exemplified by Jurassic pliosaurs, such as Pliosaurus and Liopleurodon (Figure 2C) and the giant Kronosaurus from the Early Cretaceous of Australia (Figure 3), but evolved repeatedly during plesiosaur history, both in the Jurassic and the Cretaceous. Some pliosaur skulls approach 2.4 meters in length, and their teeth are among the largest marine amniote teeth to evolve. Pliosaurs continued into the Late Cretaceous with smaller heads adapted to feeding on large fish, a diet also inferred for the polycotylids. In general, plesiosaurs show little evolutionary change across the Jurassic–Cretaceous boundary and continued to thrive, both the short-necked macropredators and the long-necked fish-eaters (piscivores) (Figure 3). The Late Cretaceous also saw their further specialization, an ever-increasing neck length in the elasmosaurs (Figure 4A). Neck lengthening evolved by addition of vertebrae, not by elongation of the neck vertebrae (as in giraffes and sauropod dinosaurs). As a result, some elasmosaurs have more than 75 neck vertebrae, and the neck of the eponymous Elasmosaurus from the Late Cretaceous Western Interior Seaway of North America is seven meters long! While mainly being piscivores, some elasmosaurs appear to have been filter-feeding. Plesiosaurs still thrived in the latest Cretaceous (Figure 3), as documented in Antarctica, but all fell victim to the Cretaceous–Paleogene mass extinction event 66 million years ago, bringing their illustrious history to an end.
Figure thumbnail gr4
Figure 4Plesiosaur biology.
Show full caption
View Large ImageFigure ViewerDownload Hi-res imageDownload (PPT)
Plesiosaur biology
Plesiosaurs were medium-sized to very large marine reptiles, ranging in total length from under two meters to over 12 meters (Figure 2). Small plesiosaurs may not have exceeded 100 kilograms in body mass and medium-sized ones a few to several hundred kilograms. Among the largest plesiosaurs, such as elasmosaurs, in which half of the length was taken up by the neck, body mass calculations yielded up to six metric tons (Figure 4C), whereas the largest pliosaurs over 10 meters long must have exceeded 15 metric tons. Unlike for dinosaurs, body mass has been little studied in plesiosaurs, and estimates are only based on volumetric calculations from life models, drawings and three-dimensional capture of mounted skeletons. No empirical relationships between any skeletal measurement, such as skull length or humerus length, and body mass have been established.
The plesiosaur skeleton (Figures 1 and 2) is well known because of many finds from sedimentary rocks laid down in poorly oxygenated quiet waters (such as the Early Jurassic deposits from the Dorset coast) preserving the entire skeleton (albeit the small head of the long-necked forms is often missing). Skull architecture is relatively uniform with a single temporal opening and little variation in suture patterns. The small-headed piscivores had short snouts (Figure 2A,D), often armed with slender recurving and interlocking teeth. Whereas their long neck intuitively looks very flexible, modern research indicates a surprisingly stiff structure based on the tight articulation of the vertebrae with each other (Figures 1 and 2). The function of the long, stiff neck probably was for visual and hydrodynamic concealment of the large body from the prey, allowing efficient harvesting of small, schooling fish from a distance.
Larger-headed forms, including the giant macropredatory ones, had a rather crocodile-like snout with large fangs and a constriction behind the anterior ‘crown’ of fangs (Figure 2C,F). Their teeth are more robust but mostly retain the basic conical shape with distinct, sometimes sharp, ridges formed by the tooth enamel. Pliosaurus had trenchant teeth with a triangular cross section and two sharp cutting edges for dismembering large prey items, including ichthyosaurs and other plesiosaurs but also sea turtles. Predation of this kind is documented by fossilized plesiosaur stomach contents.
The plesiosaur trunk was short and rigid, with roughly 20 dorsal vertebrae. The single-headed ribs together with the very well developed ‘basket’ of belly ribs define a streamlined, but wide body (Figures 2 and 4). The connection between the shoulder girdle and the vertebral column appears weak, because the scapula is small and does not overlie the rib cage (Figure 2). This is puzzling with regard to flipper function and swimming style. The ventral parts of both the shoulder girdle and pelvic girdle are large, flat plates of bone (Figures 2 and 4D) that appear mechanically linked to the gastral basket. The pelvic girdle is weakly attached to the vertebral column by a single sacral rib. The tail is short, not exceeding the length of the trunk, and rapidly tapering to a point. Whether plesiosaurs had a small tail fin is controversial.
Plesiosaur limbs were modified into stiff flippers with simplified, flattened upper and lower limb bones (Figures 2 and 4). However, unlike in ichthyosaurs, the uniformly five digits remained discrete and had a greatly increased number of phalanges. Muscle reconstruction in conjunction with functional finite element analysis indicates that the discrete fingers effected torsion of the flippers around their long axis during underwater flight (Figure 4D,E). Historical skeletal mounts and paleoart generally depict a rowing (drag-based) style of underwater locomotion, but modern consensus is four-winged underwater flight (lift-based locomotion). Clear morphological evidence for lift-based over drag-based locomotion is the shape of the flippers, long and slender and tapering to a point (Figures 2 and 4). The upstroke of the plesiosaur flipper appears to have been weaker than its downstroke because of the relatively small attachment areas available for the muscles lifting the flippers.
Such four-winged underwater flight does not exist in living animals (sea turtles, penguins, and sea lions only use their front flippers in various kinds of underwater flight). Thus, understanding how it works in plesiosaurs has been challenging — but also inspiring from a biomimetic perspective. One issue has been the coordination of the front flippers with the hind flippers, because the latter had to cope with the vortices generated by the former. Analog hydrodynamic models (Figure 4F) show that proper coordination, flipper spacing and geometry result in greatly improved thrust and efficiency of the system compared to two independent sets of flippers. In addition to the four flippers allowing faster acceleration for effective ambushing, they may have also provided greater maneuverability than two flippers. This has inspired a biomimetic four-winged underwater robot, but further analog and digital modelling of the plesiosaur four-winged underwater flight will be rewarding. In particular, one is left to wonder why marine turtles and mammals did not evolve a plesiosaur-like system of propulsion.
Preciously little is known about the plesiosaur integument despite their fossils occurring in Lagerstätten famous for their soft part preservation. Plesiosaur skin presumably lacked scales but was smooth and was underlain by a thick layer of blubber, serving both insulation and hydrodynamics. Likewise, little is known about plesiosaur sense organs. They neither have particularly large nor small eyes, and their inner ear labyrinth shows similarities with that of other pelagic amniotes. However, the snout may have harbored a special sense organ based on its rich internal vascularization. No direct evidence is available for the respiratory system, but any long-necked plesiosaur must have had a very large lung volume to compensate for dead space in their long trachea.
Several lines of recent evidence indicate that plesiosaurs were fully endothermic, hence the inferred existence of blubber as an insulating material. For one, their unique fossilized bone histology indicates extremely fast growth, and bone histomorphometry infers metabolic rates comparable to those of birds (Figure 5). Preserved molecular metabolic markers point in the same direction. Bone histology and a singular find of a gravid female (Figure 2E) inform plesiosaur life history strategies. They apparently were extreme K strategists, as a single fetus was born at 60% of the mother’s size, growing to full size within a few years (Figure 5). For comparison, this is much faster than similar-sized cetaceans and pinnipeds. Nothing is known about parental care, sexual dimorphism, social behavior or migration. There is limited evidence, again from bone histology, for deep diving. As any large pelagic animal, plesiosaurs must have been capable divers in order to harvest enough food for supporting their large size and fast metabolism. The somewhat regular occurrence of gastroliths (stomach stones) within plesiosaur skeletons was once viewed in the context of deep diving but the gastroliths more likely helped stabilize the body.
Figure thumbnail gr5
Figure 5Microanatomy and histology of plesiosaur bones in thin section.
Show full caption
View Large ImageFigure ViewerDownload Hi-res imageDownload (PPT)
Global dispersal and habitat
Analysis of plesiosaur biogeography based on their recently solidified phylogenetic tree is lacking, but similarities in plesiosaur faunas around the world suggest that these animals had a high dispersal capability, colonizing the entire globe. Dispersal was facilitated by their low-energy cruising style combined with endothermy. Plesiosaur skeletons from the Jurassic and the Cretaceous (Figure 2) are mostly found in deposits of the open sea, often together with other large marine reptiles such as ichthyosaurs and mosasaurs (in the Late Cretaceous). Commonly, such deposits also contain pterosaur fossils. A 2017 compilation recognized nearly 120 plesiosaur species.
Classical plesiosaur Lagerstätten are the lowermost Jurassic of the Dorset coast of the UK, the Jurassic Posidonia Shale of Germany, the Oxford and Kimmeridge Clays and similar deposits of the UK, France and the Arctic archipelago of Svalbard, and the Late Cretaceous Niobrara Chalk and similar deposits of the Western Interior Seaway of North America. However, plesiosaur fossils are found around the globe, often in remote places. A non-comprehensive listing includes the latest Cretaceous finds from Seymour and Vega islands, Antarctica, South American finds from Chile, Argentina and Columbia, African plesiosaurs from Angola and Morocco, plesiosaur fossils from New Zealand and Australia (sometimes preserved as precious opal), and finally Asian ones from Japan and Russia. These finds are mostly from Cretaceous rocks, primarily reflecting the much wider distribution of these younger deposits compared to the older Jurassic ones. The sedimentology and faunal composition of these Lagerstätten listed above would suggest that plesiosaurs were less pelagic than ichthyosaurs, but a test of this hypothesis is lacking. However, unlike ichthyosaurs, but like smaller cetaceans, plesiosaurs also invaded freshwater habitats, for instance in the Late Cretaceous Dinosaur Park Formation of Alberta, Canada.
The relevance of plesiosaurs
Plesiosaurs have much to offer to our understanding of the process of evolution and the history of life. This was already understood by the early geologists, paleontologists and zoologists. Cuvier had posited that the changes in life forms through time were caused by a series of catastrophes, while Darwin, inspired by Lyell’s gradualism, had emphasized the gradual transformation of species; our modern understanding of the plesiosaur reveals the truth in both views. Plesiosaurs gradually evolved their unique Bauplan, which then remained essentially unchanged for 140 million years. It went extinct not because of poor adaptation, but through catastrophic geological and cosmological processes.
Secondary marine adaptation is one of the big trends in tetrapod evolution, and one that makes convergent evolution particularly obvious. The standard model of the marine amniote, that of the tail-propelled swimmer, first arose in ichthyosaurs (Figure 1B) and early sauropterygians but then many times again during the entire Mesozoic (e.g., marine crocodiles and mosasaurs) and Cenozoic (e.g., cetaceans and sirenians). Plesiosaurs, on the other hand, remain unique, adding a novel evolutionary solution to living in the sea as an amniote (Figure 1F). Nevertheless, plesiosaurs share certain nearly universal adaptations, such as endothermy and vivipary, with other fully marine amniotes, extinct and extant. Because plesiosaurs are extinct, we have to understand them with the methods of paleontology but have to explain them with the knowledge of modern biology. Plesiosaurs are thus model organisms for evolution, and many more fundamental insights are to be expected from their continued study.
ADDITIONAL RESOURCES